Key Result
This research has identified new check lines of B. napus to use for secondary dormancy screening. The results also demonstrate the need for screening for secondary dormancy potential (SDP) in commercial seedlots.
Project Summary
Background
Canola acreage on the Canadian Prairies in 2017 totaled more than nine million hectares, with over half the hectares grown in Saskatchewan. Only about 50% of canola seed planted germinates and emerges with the other 50% remaining in the seed bank. Volunteer canola ranks 4th in most occurring weeds on the Canadian Prairies and creates a problem for control in other herbicide tolerant crops. The unintentional movement of genes between cultivars and possibly species due to pollen transfer is of great concern due to gene stacking leading to herbicide resistance build up. It is believed secondary dormancy is the physiological mechanism leading to the extended presence in the seed bank. The induction potential for secondary dormancy varies greatly among Brassica napus lines and is largely influenced by the environment and genetics.
Prior to this project, there was very limited knowledge on the range of secondary dormancy potential (SDP) across spring B. napus other than earlier studies showing many of the open-pollinated varieties of canola grown in the 1990s and early 2000s had moderate to high SDP. This study identified a greater range in SDP across spring B. napus with some lines showing consistent high or low SDP across seed production environments, and other lines that showed different SPD depending on the growth environment. Furthermore, SDP was not associated with seed vigour or germination parameters. Other results supported the decrease of SDP over time and with exposure to non-freezing or varied temperatures which indicates there is little potential commercial seedlots increase in SDP based on exposure to temperature changes during storage and cleaning. However, when commercial seedlots were tested, differences in SDP between varieties and seed production environments were also observed, but not to the extreme high range observed in the diversity collection.
Objectives
The role seed dormancy plays in establishment of canola stands on the Canadian Prairies is not clear; however, previous work has showed variation for secondary seed dormancy in Canadian canola cultivars, with more than half of the cultivars being classified as high. Furthermore, knowledge of overall seed biology and how it relates to germination, vigour of seed and seedlings and dormancy potential in B. napus are relatively poorly understood.
This project aimed to answer the following questions:
- Is there any evidence that secondary dormancy varies in commercially germplasm and how does the range compare to the diversity found across spring B. napus?
- What is the relationship between precocious germination, primary and secondary dormancy and how does the potential of these relate to seedling vigour and other seed-biology related traits? Based on these results, could selection for reduction in secondary dormancy have adverse effects on stand establishment traits?
- Is there any evidence that seed storage, conditioning treatments inducing secondary dormancy in HDP lines?
- How heritable is dormancy and seedling vigour-related traits and how complicated are the underlying genetics? How feasible is it for public and private canola breeders to select for or against dormancy, seed and seedling vigour and are there molecular markers associated with genomic regions contributing to these traits?
- What role do the major seed storage proteins cruciferin and napin play in secondary dormancy expression? Is the selection for low glucosinolates lines unintentionally selecting for higher dormancy potential expression due to the linkage of glucosinolates and napin genes?
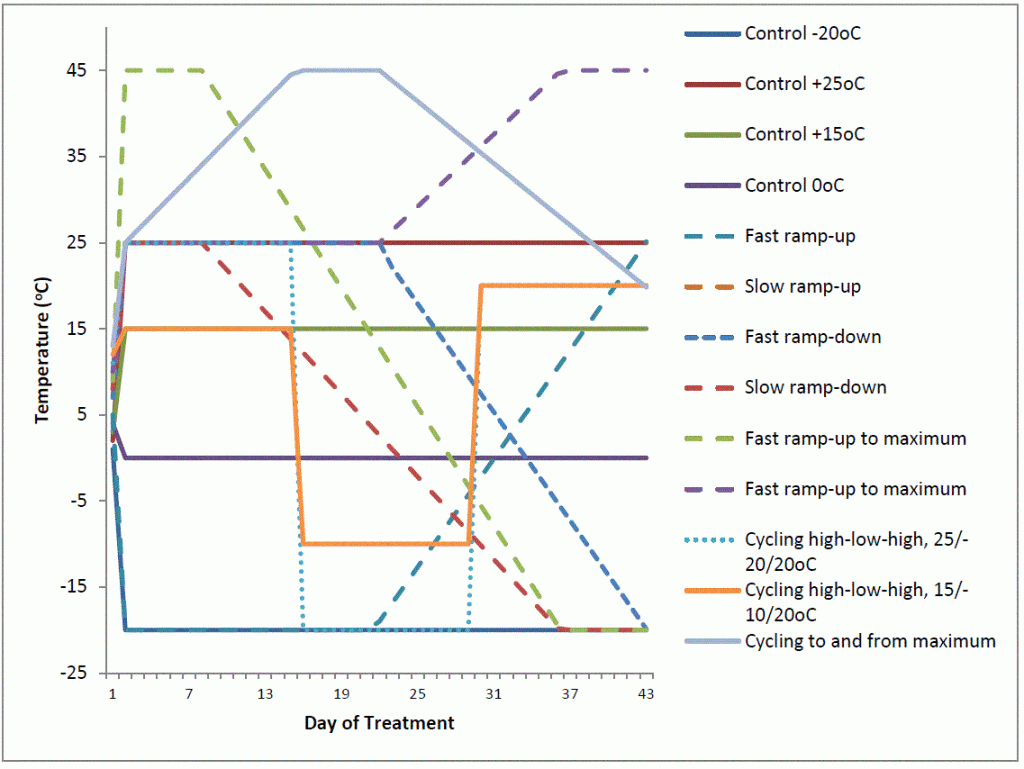
Results
Results from this project show a very wide range in SDP across a spring B. napus diversity collection with several high SDP lines being much greater than those identified in the earlier studies. Additionally, the effect of seed production sites was evident, especially for a subset of diverse lines which exhibited contrasting SDP between sites. The SDP values for all seedlots have been compared to total germination, rates of germination, primary dormancy, seed vigour measurements (controlled deterioration test and pre-chill germination test) seed size and precocious germination. No association between SDP and any of these other germination or seed vigour parameters have been found. Thus, the results indicated that SDP is independent of routinely employed assessments or measurements of seed vigour. These results also lay the groundwork to study the genetics controlling SDP.
In order to test the relevance of the findings to the commercial hybrid seed production industry, the researchers initiated two smaller studies looking at i) the effect of temperature regimes that commercial hybrid seed is exposed to after harvest until the point of sale to the grower on SDP; and ii) SDP of commercial seedlots from different seed production environments. Examination of 12 extreme temperature regimes imposed on seed of high and low SDP lines confirmed that SDP decreases over time and with exposure to non-freezing temperatures and that none of the consistent or varied temperature regimes increased the SDP of seedlots. Examination of 30 commercial seedlots of hybrid canola have confirmed that variation within breeding programs exists for SDP and that for high SDP varieties, seed production environment contributes to significant increases of SDP for a given seedlot. These results will be of value for the canola seed production industry in prioritizing selection for decreased SDP in commercial varieties and could have implications on seedlot blending and/or seeding rate recommendations.
Conclusions
This research has identified new check lines of B. napus to use for secondary dormancy screening. Previously, little was known of consistent high/low checks other than results reported in the early 2000s. By screening a diversity collection that was produced in more than four environments, the researchers established a collection of lines (6 in total) that produce consistently high SDP and contrasting low SDP regardless their maternal environment. The lines with the consistently high SDP phenotypes are much larger and more consistent than lines previously reported. The researchers anticipate providing seed of these lines for use as checks in commercial seed testing laboratories and to hybrid production companies.
These results also demonstrate the need for screening for secondary dormancy potential (SDP) in commercial seedlots. Currently, standard germination and seed vigour testing are routinely performed. The researchers tested these ‘standard’ protocols and compared them to screening for SDP on the same seedlots and found the results of the standard protocols were all highly correlated or predicted results of a parallel protocol; however, the SDP screening was not related to the standard protocols. Furthermore, it was demonstrated that in some commercial seedlots, a significant portion of the seed has potential to be induced into secondary dormancy. The researchers are working with hybrid seed producers and commercial seed testing facilities to re-examine seeding rate and blending recommendations based on these results.